

1. Introduction
In this study, Santan et al. conducted an experiment on the use of controlled illumination to improve vision in fishes for effective predator detection. Prior to their study, cases of controlled illumination were considered rare and only limited to chemiluminescence, which only functions in darkness. However, the studies on Tripterigion delaisi, a small triplefin, revealed that they can reflect the sunlight sideways through their iris, and prompted the authors to test if ‘diurnal active photolocation’ also improves T. delaisi’s ability to detect the cryptobenthic sit and wait predator, Scorpaena porcus, a scorpion fish with strong daytime retroreflective eyes .
The retroreflective eyes of scorpionfish improve its camouflage by concealing its black pupils, and make its detection difficult. Therefore, diurnal active photolocation can be called as an adaptation feature of triplefins to detect their predator with the help of their ocular sparks. An ocular spark is a radiance produced on the iris of the triplefin when its lens (which protrudes from the pupil) allows some of the downwelling light to pass through it without entering the eyes and focuses those rays to form a bright spot on the iris below. The iris then reflects those rays sideways helping the triplefin to detect a nearby scorpionfish by increasing the contrast of the eyes of the scorpionfish.
In the paper, the authors present the three different independent experiments where they manipulated the ocular spark production in triplefins, and compared their response to the control visual stimulus— a scorpionfish, or a stone—using three different treatments: without using any hat, using a clear mini-hat, and using an opaque mini-hat, over triplefins’ head. The authors predicted the triplefins with unhatted and clear hatted control to keep a greater safe distance from the scorpionfish than opaque-hatted triplefins.
2. Materials and Methods
-
Model Species
The authors used Tripterigion delaisi, triplefins having standard length of 3–5 cm, and scorpionfish Scorpaena porcus, a cryptobenthic sit and wait predator (12-20 cm) for the experiment. Whereas the eyes of the scorpionfish contain reflective stratum argenteum and translucent retinal pigment epithelium contributing to its diurnal eyeshine, triplefins have high visual acuity and contrast sensitivity to effectively sense any change in scorpionfish’s pupil.
-
Mini Hats for Control Treatment
The researchers anesthetized the triplefins for 5-10 minutes, and separately glued either mini opaque hats (shading hat treatment) made up of dark red polyester filter or mini transparent hats (clear-hatted control) on dorso-posterior head area of the triplefins by using surgical glue. They didn’t attach any hat but followed the similar procedure in other triplefins (unhatted sham control). Clear-hatted and unhatted triplefins regularly produced the ocular sparks (fig.1a,b), whereas the sparks were absent in shading hatted triplefins(fig1c). Hence, the treatments were successful in producing the desired effects.
-
Laboratory Experiment
They used four aquaria (L x W x D: 130 x 50 x 50 cm3) and placed perforated containers (L x W x H: 24 x 14 x 16 cm3) at the end of each aquaria where scorpionfish was placed on two and stone on others. The containers with stimuli were shaded. On day 1, unhatted, hatted and clear hatted triplefins were placed in the aquaria, and allowed time to explore the new environment. The motive was to note the distance of each triplefin from the stimulus. On day 2, observation was taken with scorpionfish visible on two aquaria and stone on others. On day 3, stimuli were swapped and observations were recorded again. Although 20 triplets (n=60) were used, only 15 triplets were used for analysis owing to premature hat loss in five others.

-
First Field Experiment
The researchers replicated the laboratory experiment by using 10 Evotron Plexiglas tanks (L x W x D: 130 x 25 x 50 cm3) placed at 15 m depth on a sandy patch, with five facing north and five facing south as a control for the position of sun (fig.1g,h). At the end of the triplefin compartment (L x W x D: 125 x 25 cm2) was the stimulus display compartment (L x W x D: 15 x 25 cm2), and a similar sized compartment behind the latter was used to hide the alternative stimulus. The distance of individual triplefins from the stimulus was noted as the main response variable. First set of observations were taken the next day after the release of triplefins in the compartment. Stimuli were swapped for second observation day. Fifty triplets were tested, twenty-five per orientation (north or south facing).
-
Second Field Experiment (Immediate Response)
After the researchers discovered that the recovery from anesthesia is completed in 2 – 3 hours, shorter tanks are sufficient to reveal a response, and mobility and response is faster when individuals are placed on dark sand, they realized that they could collect the data in the shorter observation time window. Keeping in mind these new modifications, they set up a layout similar to the first field experiment with a slight variation in container dimensions: 50 x 25 cm2 for triplefins and 12 x 25 cm2 for scorpionfish. One triplefin was released to each of the 10 tanks at a time, and all the data concerning their immediate response was completely taken within 90 – 100 minutes—divided into three 30 min interval. In total, 38 shaded and 42 clear-hatted triplefins were observed.
-
Statistical Analysis
Statistical computing was done in R. In the laboratory, researchers made the virtually error-free measurements at ±1 cm scale, as the observer, fish, and ruler were close to each other and easy to align. However, for the field experiment, repeatability analysis was necessary. Two SCUBA divers noted the 116 distances of triplefins in the first field experiment, and the repeatability estimate was calculated to be 0.995 using rptR package with data type Gaussian and 1000 permutations (likelihood ratio test: p < 0.0001).
The response variable, i.e. the distance from the display compartment, was in two linear and one generalized linear mixed models (GLMMs). The researchers compared the two control treatments (unhatted and clear hatted) for the first two experiments to confirm that carrying a hat did not affect triplefins’ general behaviors and their ability to distinguish a cryptic predator from a stone.
In the analysis of the first field experiment (fig.2b,c), we can see that there is the variation in triplets number. This is because the response of clear hatted and unhatted were averaged (owing to their similarity) to recover some data points lost as the result of hat loss: only 22 out of 50 tested triplets had remained intact.
-
Visual Modeling
The experimental results that showed ocular sparks helping triplefins to detect scorpionfish was necessary to validate. It is where the researchers used visual modeling. They modeled the effect of the blue ocular spark (stronger out of two known for T. delaisi). Using retinal quantum catch estimates, they calculated the chromatic contrast between the scorpionfish pupil with and without the contribution of ocular sparks. For the same two conditions, they also estimated the achromatic contrast by calculating Michelson contrast, i.e.
C = (Q1 – Q2) / (Q1 + Q2)
where Q1 and Q2 are the sum of quantum catches of the two double cone photoreceptors of T. delaisi. They also calculated the maximum discernible distance of ocular spark’s reflection from scorpionfish pupil.
3. Results

-
Long term Response
Response observed in the laboratory, and the first field experiment is the long-term response since enough time (overnight) was provided for acclimatization. The researchers noted that triplefins maintained a greater distance from the scorpionfish than the stone irrespective of the hat treatment which pointed out the fact that light-redirection has no effect to visually distinguish scorpionfish from stone. While the shaded triplefins stayed significantly closer to scorpionfish as compared to unhatted and clear hatted, this was not the case for the stone. Remarkably, the effect of hat treatment was only present in laboratory and north-facing tanks, and absent in the south-facing tanks.
-
Immediate Response
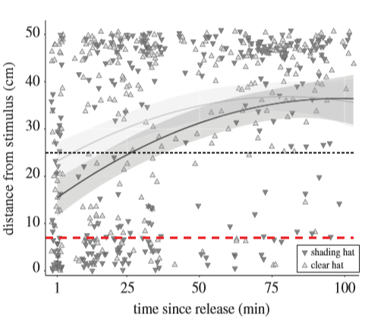
Response observed for the second field experiment is the immediate response since no acclimatization time was provided. Many triplefins, after being released to the tank, moved towards scorpionfish, presumably being attracted by the shaded display compartment. The researchers calculated that, after 1 minute, 47% of the shaded triplefins, compared to only 21% of clear hatted triplefins, had come closer than 7cm (dotted red line in fig.3), which is a distance at which active photolocation with unobstructed ocular sparks is predicted to occur. Clear-hatted triplefins retreated to opposite end 20 min earlier than shading-hatted, and north-south orientation had no effect.
-
Visual Modeling

Results from the visual modeling showed that the ocular sparks could not produce any discriminable chromatic contrast at any distance between the triplefin and the scorpionfish. However, achromatic Michelson contrast was easily detected across a broad range. The researchers found that the reflections of the ocular spark from the scorpionfish pupil was detectable up to 6 cm on average conditions for north facing triplefins, which increased by 2 – 3 cm for south-facing triplefins. The average detection range of 6-8 cm matches with the 7 cm threshold shown in figure 3.
4. Conclusion
All of the three experiments showed that the triplefins kept a smaller distance from a well camouflaged scorpionfish when their ocular sparks are suppressed by opaque hats. With this, the authors were successful in giving the first proof of the concept of diurnal active photolocation of the cryptobenthic predator. However, diurnal active photolocation is not something that is strictly required by triplefins to detect the presence of scorpion fish. Triplefins were able to maintain a greater distance from the scorpionfish than from the stone regardless of their hatting treatment. Therefore, diurnal active photolocation is only the mechanism that supplements triplefins’ ability to detect their predator.
Reference (The Original Paper)
- Santon Matteo, Bitton Pierre-Paul, Dehm Jasha, Fritsch Roland, Harant Ulrike K, Anthes Nils, Michiels Nico K. 2020 Redirection of ambient light improves predator detection in a diurnal fish Proc. R. Soc. B.28720192292
Was this summary helpful?
Mention in the comments section below.
Do not forget to check other research paper summaries and general blogs.







